Why should mitochondria define species?
Stoeckle and Thaler 2018
A Critique
|
For he who is acquainted with the paths of nature, will more readily observe her deviations; and vice versa, he who has learnt her deviations, will be able more accurately to describe her paths.
—Francis Bacon, Novum Organum
|
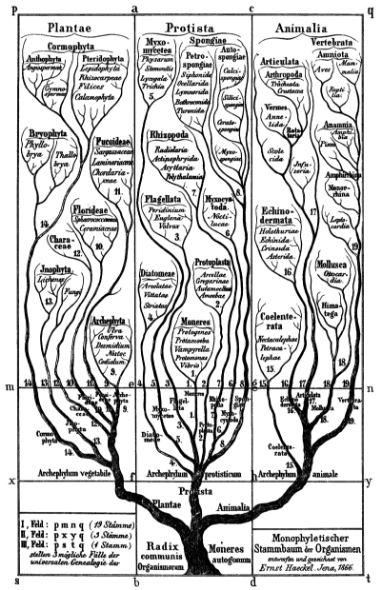 Ernst Haeckel’s tree of life (Generelle Morphologie der Organismen, 1866). This is the topology pictured by those who assume evolution can be reasonably described in terms of trees of descent rather than webs.
Ernst Haeckel’s tree of life (Generelle Morphologie der Organismen, 1866). This is the topology pictured by those who assume evolution can be reasonably described in terms of trees of descent rather than webs.
Sorry not to get back to you sooner, but I’ve been working rather obsessively on what my new employer refers to as the “work product.” But I decided I really had to carve out some time this morning and get back to you about that Stoeckle and Thaler paper you asked about.
I have to admit that, at the very outset of reading through their paper, I raised my eyebrows. They start with the statement that there is a high level of correspondence between barcodes and “species” even though the word species itself has no clear meaning. Indeed, what they refer to as a “consensus of domain experts” seems to reflect mere belief on Stoeckle and Thaler’s part rather than an actuality. Certainly they must be relying on belief, because they cannot possibly have actually verified that there is consensus on the classification of organisms, for, as they themselves say, there are millions of them. Moreover, the fact that classifications at every taxonomic level continue every year to be revised suggests that this consensus is a figment. So I think their statements just reflect their faith that these “domain experts” have done their work correctly.
 Biologists who still think in terms of strictly bifurcating trees of life are stuck in a mindset that hasn’t changed in any fundamental way since the days of Ernst Haeckel. And yet, a huge amount of data now demonstrates that hybridization is pervasive.
Biologists who still think in terms of strictly bifurcating trees of life are stuck in a mindset that hasn’t changed in any fundamental way since the days of Ernst Haeckel. And yet, a huge amount of data now demonstrates that hybridization is pervasive.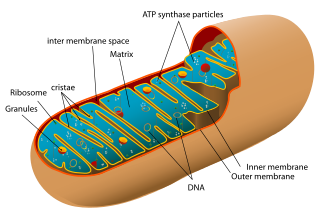 Schematic of a mitochondrion. Mitochondria contain only about 1% of the DNA in a typical eukaryotic cell, the rest is in the nucleus.
Schematic of a mitochondrion. Mitochondria contain only about 1% of the DNA in a typical eukaryotic cell, the rest is in the nucleus.
To anyone, like me, who thinks in terms of pervasive hybridization during the course of evolution, estimated rates of mutation in “barcode” (mitochondrial) sequences are highly suspect. Accepted notions about the magnitude of such rates are based on studies that assume evolution can be reasonably described in terms of trees of descent rather than webs. If that assumption is wrong, so are all the estimates.
Under the perspective that assumes hybridization has been pervasive, the various stable populations existing in nature each have a characteristic barcode because the stabilization processes that initially create them are typically local events affecting few individuals. Indeed, the initial event can in some cases be a single female crossing with a single male, in which case, a single mitochondrial haplotype (barcode) is inherited from the mother (mtDNA is strictly maternally inherited). So all members of the newly stabilized population, which would all be descended from her, would also have that haplotype.
Even in those cases where multiple females play the role of mothers in more complex stabilization processes, certain factors would tend to reduce the variation of the haplotype that could be contributed. For example, hybrid zones are geographically localized, so that potential haplotypes that can be passed to hybrid offspring come only from that part of the maternal population adjacent to the zone, which might greatly reduce the potential variation that could be sampled in comparison with that present in the maternal population as a whole.
An entirely different factor that can reduce mtDNA haplotype variation is stabilization itself. Computer simulations of highly variable hybrid populations indicate that stabilization processes initiate when two individuals with the same fully paired karyotype happen to mate with each other. This is because (1) they alone will produce offspring that are karyotypically identical to themselves, (2) their fully paired karyotype, which greatly increases the production of functional gametes, allows them to produce those offspring in much larger numbers than can other hybrids in the population who lack full pairing, and (3) those many offspring with fully paired karyotypes, too, will be far more fertile (than others who lack full pairing) and will be spatially localized (in the vicinity of where their parents produced them), so they, will tend to mate with each other and due to their higher fertility produce many offspring. In this way a spatially localized dense kernel of founding individuals with high fertility is created. But note that all individuals descended from this stabilization process will share the haplotype of the founding female who initiated the stabilization process. So, again, due to this factor, the variation in haplotype will be very low. In fact, nonexistant.
To this tendency of stabilization processes to reduce mitochondrial diversity, add stabilization theory’s emphasis on the extreme accuracy of DNA replication (as opposed to Darwinian theory’s emphasis on the sufficiency of mutation rates to produce observed evolutionary changes) and it becomes obvious that most populations, even those founded long ago, will evince very low levels of mtDNA variation.
However, Stoeckle and Thaler (p. 23), thinking inside a box built of boards sawn from the tree of life, propose what they call a simple hypothesis, i.e.,
In other words, they are saying that because
- biologists have concluded, on the basis of observed levels of human mitochondrial variation, that the common ancestor of all human mtDNA dates to about 200,000 years ago;
and - since human mitochondrial variation is of about the same magnitude as the mitochondrial variation typically seen in other kinds of organisms,
 My book on hybridization in birds (Handbook of Avian Hybrids, Oxford U. P., 2006) documents more than 4,000 hybrid crosses among in birds, about half of which occur in a natural setting, and lists hundreds of hybrids that have been treated as species.
My book on hybridization in birds (Handbook of Avian Hybrids, Oxford U. P., 2006) documents more than 4,000 hybrid crosses among in birds, about half of which occur in a natural setting, and lists hundreds of hybrids that have been treated as species.
we can conclude that “essentially all other animal species” also “expanded from mitochondrial uniformity within the past 200,000 years.” In the news, this mere hypothesis has been trumpeted as a huge revelation, for example the headline, “Massive Genetic Study Reveals 90 Percent Of Earth’s Animals Appeared At The Same Time!”
But one can just as easily interpret the observed pervasively low levels of mitochondrial diversity as evidence that new forms of life typically come into being via stabilization processes (as proposed by stabilization theory), which truly is a simple hypothesis. Any population founded by a stabilization process is expected to have low mitochondrial diversity because such processes constitute genetic bottlenecks, that is, the number of founders involved in the founding of such populations is typically very small.
Stoeckle and Thaler have formulated their reasoning in terms of the tree outlook. So their conclusions stand or fall just as the correctness of that outlook stands or falls. The woodchoppers of biology have been hacking away at that particular tree for a long, long time. Every plant or animal of hybrid origin is another gash in the trunk. Soon, it just may topple over.
If you have questions on a biological topic, feel free to contact the author.
Stabilization theory provides an intellectual basis for Charlie Chaplin's dream: